Las poblaciones viven como parte de una comunidad -un conjunto de organismos distintos que habitan un ambiente común y que están en interacción recíproca-. Se reconocen tres tipos principales de interacción específica en las comunidades: la competencia, la depredación y la simbiosis. Cuanto más semejantes sean los organismos en sus requisitos y estilos de vida, más probable es que la competencia entre ellos sea intensa. Como resultado de la competencia, la aptitud total de los individuos que interactúan puede verse reducida.
La importancia relativa de la competencia sobre la composición y la estructura de la comunidad es actualmente una cuestión de debate entre los ecólogos, al igual que los métodos utilizados para estudiar la competencia y otros aspectos de la ecología de la comunidad. La depredación es la alimentación con organismos vivos; tiene una fuerte influencia sobre la evolución delas poblaciones, tanto de predadores como de presas.
Las interacciones depredador-presa también influyen sobre la dinámica de la población y pueden aumentar la diversidad de las especies reduciendo la competencia entre las presas. La simbiosis es una asociación íntima y a largo plazo entre organismos de especies diferentes. Puede ser beneficiosa para una especie e indistinta para la otra (comensalismo), beneficiosa para una y perjudicial para la otra (parasitismo), o beneficiosa para ambas especies (mutualismo).
Quedan por responder dos preguntas importantes en lo que hace a la composición de la comunidad y a su estructura: ¿Qué determina el número de especies en una comunidad?, ¿qué factores sustentan los cambios en la composición de la comunidad con el transcurso del tiempo?
Dado su tamaño y su aislamiento relativo, las islas pequeñas suelen ser excelentes laboratorios naturales para el estudio de los procesos evolutivos y ecológicos. De acuerdo con la hipótesis de la biogeografía de islas, el número de especies en las islas alcanza un equilibrio determinado por el equilibrio entre la inmigración y la extinción.
De acuerdo con la hipótesis de la perturbación intermedia, la mayor diversidad de especies es hallada en comunidades como las selvas lluviosas tropicales y los arrecifes de coral, sometidas a perturbaciones ambientales a una frecuencia intermedia. Las comunidades en las cuales las perturbaciones son infrecuentes o muy frecuentes, generalmente tienen una diversidad de especies más baja. Después de las perturbaciones ambientales, las comunidades se recolonizan por la dispersión de formas inmaduras que provienen de comunidades vecinas.
Si transcurre suficiente tiempo antes de la siguiente perturbación importante, una comunidad pasa generalmente a través de un proceso de sucesión ecológica en el que los primeros colonizadores son remplazados por otras especies, que a su vez pueden ser remplazadas por otras más. La evidencia actual indica que las comunidades, al igual que las poblaciones de las cuales están compuestas, son dinámicas, y cambian continuamente a medida que cambian las condiciones.
La importancia relativa de la competencia sobre la composición y la estructura de la comunidad es actualmente una cuestión de debate entre los ecólogos, al igual que los métodos utilizados para estudiar la competencia y otros aspectos de la ecología de la comunidad. La depredación es la alimentación con organismos vivos; tiene una fuerte influencia sobre la evolución delas poblaciones, tanto de predadores como de presas.
Las interacciones depredador-presa también influyen sobre la dinámica de la población y pueden aumentar la diversidad de las especies reduciendo la competencia entre las presas. La simbiosis es una asociación íntima y a largo plazo entre organismos de especies diferentes. Puede ser beneficiosa para una especie e indistinta para la otra (comensalismo), beneficiosa para una y perjudicial para la otra (parasitismo), o beneficiosa para ambas especies (mutualismo).
Quedan por responder dos preguntas importantes en lo que hace a la composición de la comunidad y a su estructura: ¿Qué determina el número de especies en una comunidad?, ¿qué factores sustentan los cambios en la composición de la comunidad con el transcurso del tiempo?
Dado su tamaño y su aislamiento relativo, las islas pequeñas suelen ser excelentes laboratorios naturales para el estudio de los procesos evolutivos y ecológicos. De acuerdo con la hipótesis de la biogeografía de islas, el número de especies en las islas alcanza un equilibrio determinado por el equilibrio entre la inmigración y la extinción.
De acuerdo con la hipótesis de la perturbación intermedia, la mayor diversidad de especies es hallada en comunidades como las selvas lluviosas tropicales y los arrecifes de coral, sometidas a perturbaciones ambientales a una frecuencia intermedia. Las comunidades en las cuales las perturbaciones son infrecuentes o muy frecuentes, generalmente tienen una diversidad de especies más baja. Después de las perturbaciones ambientales, las comunidades se recolonizan por la dispersión de formas inmaduras que provienen de comunidades vecinas.
Si transcurre suficiente tiempo antes de la siguiente perturbación importante, una comunidad pasa generalmente a través de un proceso de sucesión ecológica en el que los primeros colonizadores son remplazados por otras especies, que a su vez pueden ser remplazadas por otras más. La evidencia actual indica que las comunidades, al igual que las poblaciones de las cuales están compuestas, son dinámicas, y cambian continuamente a medida que cambian las condiciones.
Competencia
La competencia es la interacción entre individuos de la misma especie (competencia intraespecífica) o de especies diferentes (competencia interespecífica) que utilizan el mismo recurso; éste suele estar en cantidad limitada. Como resultado de la competencia, el éxito biológico -o sea, el éxito en la reproducción- de los individuos que interactúan puede verse reducido. Entre los muchos recursos por los cuales los organismos pueden competir se encuentran el alimento, el agua, la luz, el espacio vital, los sitios de nidificación o las madrigueras.
La competencia puede ser por interferencia o por explotación. Durante muchos años, la competencia ha sido invocada como una fuerza primordial en la determinación de la composición y estructura de las comunidades -o sea, el número y tipo de especies presentes y su distribución espacial y temporal dentro de la comunidad-. Sin embargo, recientemente diversos ecólogos han cuestionado la importancia de la competencia sobre la composición y la estructura de la comunidad. El debate -a veces cáustico- que se generó, concierne no sólo al papel de la competencia sino a los métodos que deben usarse para someter a prueba las hipótesis ecológicas. Los investigadores que echaron las bases de la ecología moderna fueron atentos observadores de la naturaleza.
Muchos de estos ecólogos creían, como tantos de sus continuadores, que la observación cuidadosa de los patrones naturales de distribución y abundancia de los organismos puede –sin manipulación experimental- producir los datos requeridos para someter a prueba las hipótesis sobre el papel que desempeñan la competencia y otras fuerzas para conformar la estructura de la comunidad. Para estos ecólogos, los patrones son los que hablan por sí mismos. Otros ecólogos sostienen que los análisis experimentales en los cuales sólo se manipula una variable por vez son los únicos medios confiables para identificar los procesos que están ocurriendo realmente en la naturaleza. En 1934, el biólogo ruso G. F. Gause formuló lo que se conoció como el principio de la exclusión competitiva.
La competencia puede ser por interferencia o por explotación. Durante muchos años, la competencia ha sido invocada como una fuerza primordial en la determinación de la composición y estructura de las comunidades -o sea, el número y tipo de especies presentes y su distribución espacial y temporal dentro de la comunidad-. Sin embargo, recientemente diversos ecólogos han cuestionado la importancia de la competencia sobre la composición y la estructura de la comunidad. El debate -a veces cáustico- que se generó, concierne no sólo al papel de la competencia sino a los métodos que deben usarse para someter a prueba las hipótesis ecológicas. Los investigadores que echaron las bases de la ecología moderna fueron atentos observadores de la naturaleza.
Muchos de estos ecólogos creían, como tantos de sus continuadores, que la observación cuidadosa de los patrones naturales de distribución y abundancia de los organismos puede –sin manipulación experimental- producir los datos requeridos para someter a prueba las hipótesis sobre el papel que desempeñan la competencia y otras fuerzas para conformar la estructura de la comunidad. Para estos ecólogos, los patrones son los que hablan por sí mismos. Otros ecólogos sostienen que los análisis experimentales en los cuales sólo se manipula una variable por vez son los únicos medios confiables para identificar los procesos que están ocurriendo realmente en la naturaleza. En 1934, el biólogo ruso G. F. Gause formuló lo que se conoció como el principio de la exclusión competitiva.
Experimentos de Gause con dos especies de Paramecium.

Los resultados de los experimentos de Gause con dos especies de Paramecium demostraron el principio de que si dos especies se encuentran en competencia directa por el mismo recurso limitado -en este caso alimento- una elimina a la otra. Paramecium caudatum y Paramecium aurelia fueron cultivados primero separadamente en condiciones controladas y con un constante suministro de alimento. Como puede verse, P. aurelia creció mucho más rápidamente que P. caudatum, indicando que P. aurelia utiliza los suministros de alimentos disponibles de un modo más eficiente. Cuando los dos protistas fueron cultivados juntos, la especie que crecía más rápidamente superó en números y eliminó a la especie que crecía más lentamente.
Un experimento con dos especies de lentejas de agua, diminutas angiospermas que se encuentran en estanques y lagos.

Una especie, Lemna polyrrhiza crece más rápidamente en un cultivo puro que la otra especie, Lemna gibba. Cuando son cultivadas juntas, Lemna gibba, que tiene diminutos sacos aéreos que la hacen flotaren la superficie, cubre a la otra especie y triunfa en la competencia por la luz. El principio de exclusión competitiva de Gause nos llevaría a pensar que sólo podrían hallarse especies disímiles coexistiendo en las comunidades naturales. Pero, en realidad, es frecuente encontrar especies ecológicamente similares que viven juntas en la misma comunidad. Esta observación planteó la cuestión del grado de similitud que debe existir entre dos o más especies para que puedan continuar coexistiendo en el mismo lugar y al mismo tiempo. Esto llevó, a su vez, al concepto de nicho ecológico. Los análisis de situaciones en las cuales coexisten especies similares han demostrado que los recursos frecuentemente están subdivididos, o son distribuidos, por las especies coexistentes.
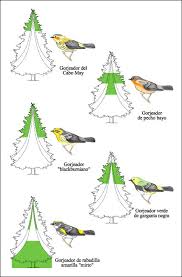
Las zonas de alimentación de cinco especies de gorjeadores norteamericanos en un abeto.
Las áreas coloreadas en el árbol indican en qué lugar cada especie pasa, al menos, la mitad de su tiempo de alimentación. Esta distribución de recursos permite que las cinco especies se alimenten en los mismos árboles. El solapamiento de nichos describe la situación en la cual los miembros de más de una especie utilizan el mismo recurso escaso. En las comunidades en las cuales ocurre solapamiento en nichos, la selección natural puede dar como resultado un aumento de las diferencias entre las especies que compiten, un fenómeno conocido como desplazamiento de caracteres.
Uno de los ejemplos de desplazamiento de caracteres citado con más frecuencia es el del pico de los pinzones de Darwin
Uno de los ejemplos de desplazamiento de caracteres citado con más frecuencia es el del pico de los pinzones de Darwin
Tamaños de los picos en tres especies de pinzones terrestres de las Islas Galápagos.

Dimensión de un nicho ecológico.
Otros ecólogos, en cambio, señalan que es imposible determinar si los distintos tamaños de pico son el resultado de interacciones competitivas que ocurrieron en épocas en que las diferentes especies coexistían en las mismas islas, o si son el resultado de adaptaciones a las condiciones locales de épocas en que las especies se encontraban aisladas unas de otras en islas diferentes. Algunos ecólogos creen que la única evidencia indiscutible del desplazamiento de caracteres consistiría en demostrar que las especies en cuestión están divergiendo actualmente en las áreas donde se las encuentra juntas.
Ambos grupos de ecólogos coinciden, sin embargo, en que cualquiera sea la causa evolutiva, las diferencias en el tamaño y la configuración del pico capacitan a las diferentes especies de pinzones para explotar diferentes fuentes de alimento y por consiguiente, para coexistir. Prácticamente, todos los ecólogos coinciden en que la competencia ocurre realmente en la naturaleza con una intensidad que varía de acuerdo con la especie implicada, el tamaño de las poblaciones que interactúan y la abundancia o escasez de los recursos. Aunque el análisis de los patrones de distribución espacial y de recursos pueden dar idea de la existencia y la importancia de la competencia en una situación determinada, se necesitan experimentos que produzcan cambios en la composición de la comunidad para demostrar que realmente hay competencia.
Competencia interespecífica entre Semibalanus y Chthamalus.
Las larvas de ambas especies se establecen en una amplia área pero los adultos viven en áreas precisamente restringidas. Los límites superiores del área de Semibalanus están determinados por factores físicos tales como la desecación. Los percebes Chthamalus, sin embargo, no viven en el área de Semibalanus, no por razones físicas (probablemente porque el área físicamente es menos limitante), sino por causa de los percebes Semibalanus. El Semibalanus crece más rápido y dondequiera que se encuentre con Chthamalus dentro de su propia área, o lo despega de las rocas, o crece por encima de él.
Este estudio y otros análogos han generado los conceptos de nicho fundamental y nicho real. El nicho fundamental describe los límites fisiológicos de tolerancia del organismo; es el nicho ocupado por un organismo en ausencia de interacciones con otros organismos. El nicho real es aquella porción del nicho fundamental realmente ocupada; está determinada no sólo por los factores físicos sino también por las interacciones con otros organismos. La mayoría de los estudios de competencia han puesto de relieve las adaptaciones y distribuciones de recursos que posibilitan la coexistencia de especies similares dentro de una comunidad. Sin embargo, ésta es una opinión sesgada, dado que es difícil estudiar las interacciones entre las especies después que uno de los protagonistas ha emigrado. Así como la competencia dentro de las especies lleva a la eliminación de la gran mayoría de los individuos, la competencia entre las especies puede llevar a la eliminación de una especie de la comunidad.
Depredación
La depredación es la ingestión de organismos vivos, incluyendo plantas por animales, animales por animales. También se considera como depredación la digestión de pequeños animales por plantas carnívoras o por hongos. Los predadores utilizan una variedad de "tácticas" para obtener su alimento. Estas tácticas están bajo intensa presión selectiva y es probable que aquellos individuos que obtienen el alimento más eficientemente, dejen la mayor cantidad de descendencia.
Mirándolo del lado de la presa potencial, es probable que aquellos individuos que tienen más éxito en evitar la depredación dejen la mayor cantidad de descendencia. Así, la depredación afecta a la evolución tanto del depredador como de la presa. También afecta al número de organismos de una población y a la diversidad de especies dentro de una comunidad.
Mirándolo del lado de la presa potencial, es probable que aquellos individuos que tienen más éxito en evitar la depredación dejen la mayor cantidad de descendencia. Así, la depredación afecta a la evolución tanto del depredador como de la presa. También afecta al número de organismos de una población y a la diversidad de especies dentro de una comunidad.
El ejemplo del lince y del "conejo" de la nieve (en realidad, una liebre).
El lince alcanza un pico de población cada 9 ó 10 años y estos picos son seguidos en cada caso por varios años de marcada disminución. Los "conejos" de la nieve, siguen un ciclo similar, con un pico de abundancia que generalmente precede al del lince en uno o más años. El tamaño de una población de predadores frecuentemente está limitado por la disponibilidad de presas. Sin embargo, la depredación no es necesariamente el factor principal en la regulación del tamaño de la población de organismos presa, que puede verse más influenciada por su propio suministro de alimentos.
Simbiosis
La simbiosis es una asociación íntima y a largo plazo entre organismos de dos especies diferentes. Las relaciones simbióticas prolongadas pueden dar como resultado cambios evolutivos profundos en los organismos que intervienen, como en el caso de los líquenes, una de las simbiosis más antiguas y ecológicamente más exitosas. Se considera generalmente que existen tres tipos de relaciones simbióticas: el parasitismo, el mutualismo y el comensalismo.
ALGUNOS EJEMPLOS DE MUTUALISMO:
Anémonas de mar en la parte posterior de una concha de caracol, ocupada por un cangrejo ermitaño.
Las anémonas protegen al cangrejo y a su vez obtienen movilidad -y así una gama de alimentación más amplia- por su asociación con el cangrejo. Los cangrejos ermitaños, que periódicamente se mudan aconchas nuevas más grandes, logran que las anémonas se muden con ellos.

Pulgones succionando savia.
Los pulgones succionan savia del floema, removiendo ciertos aminoácidos, azúcares y otros nutrientes, y excretando el resto como una melaza, o azúcar de "lerp". Esta melaza, en Australia, es cosechada como alimento por los aborígenes. Algunas especies de áfidos han sido "domesticadas" por algunas especies de hormigas. Estos áfidos no excretan su melaza al azar, sino sólo en respuesta a movimientos de las antenas y las patas delanteras de las hormigas. Los pulgones que intervienen en esta asociación simbiótica han perdido sus defensas naturales, incluyendo sus exoesqueletos duros, y dependen de sus hospedadores para su protección.

Los estorninos boyeros viven de las garrapatas que quitan de sus hospedadores. Un estornino boyero forma una asociación con un animal determinado, como el que se muestra en esta figura. La mayor parte de las actividades de estos pájaros, incluyendo el cortejo y el apareamiento, se llevan a cabo en el lomo de su hospedador.
Composición de la comunidad y el problema de la estabilidad
Visto desde una perspectiva global, las comunidades ecológicas frecuentemente parecen estar en equilibrio. Muchas especies subsisten durante muchas generaciones en grandes extensiones. Sin embargo, cuando las comunidades se examinan a escala local, se evidencia que ellas, al igual que las poblaciones de las cuales están compuestas, no están a menudo en un estado de equilibrio. Hay dos cuestiones que conciernen a la composición de las comunidades, que durante largo tiempo han sorprendido a los ecólogos. Primero, ¿qué determina la cantidad de especies de una comunidad? y, segundo, ¿qué factores sustentan los cambios en la composición de la comunidad con el paso del tiempo, como lo revela una observación detallada?
Las islas pequeñas suelen ser excelentes laboratorios naturales para el estudio de los procesos evolutivos y ecológicos debido a su tamaño y su aislamiento relativo. Los investigadores norteamericanos R. Mac Arthur y E. O. Wilson formularon la hipótesis de que la cantidad de especies en una isla dada permanece relativamente constante a través del tiempo, pero que esas especies están cambiando continuamente. De acuerdo con su propuesta conocida como la hipótesis del equilibrio de la biogeografía de islas, hay un equilibrio entre la tasa a la cual inmigran a una isla nuevas especies y la tasa a la cual una especie ya presente se extingue localmente. Aunque la cantidad de especies esté en equilibrio, su composición no está en equilibrio porque cuando una especie se extingue, habitualmente es reemplazada por una especie diferente.
Modelo de equilibrio de la diversidad de especies en una isla.
La tasa de inmigración disminuye a medida que más especies llegan a la isla, porque las especies existentes estarán mejor establecidas y, así, más capacitadas para competir con especies recién llegadas. La tasa de extinción se incrementa más rápido cuando hay un gran número de especies por el aumento de la competencia interespecífica. El número de equilibro entre las especies (línea negra) está determinado por la intersección de las curvas de inmigración y extinción. De acuerdo con el modelo de biogeografía de islas, las dos variables más importantes que influyen en la diversidad específica son el tamaño de la isla y su distancia a una fuente, habitualmente el continente, que pueda proveerle de colonizadores. Diferentes tipos de comunidades varían ampliamente en la cantidad y la diversidad de especies presentes.
Hasta hace poco se pensaba que la composición específica de estas comunidades era relativamente constante y se las citaba frecuentemente como ejemplos primarios de un estado de equilibrio. Se pensaba que su alta diversidad específica era una función de su estabilidad. Sin embargo, se ve ahora que su diversidad puede ser función, no de la estabilidad sino de la frecuencia y magnitud de las perturbaciones a las cuales están sujetas. Al principio, la diversidad en un área recién colonizada es baja. Sólo aquellas especies que están muy próximas al área perturbada y que están reproduciéndose en ese momento, son capaces de explorar el área recientemente disponible. Si las perturbaciones son frecuentes, la comunidad consistirá sólo en aquellas especies que pueden invadir, madurar y reproducirse antes de que ocurra la próxima perturbación. De acuerdo con la hipótesis de la perturbación intermedia, a medida que el intervalo entre las perturbaciones se incrementa, también lo hace la diversidad de especies. Las especies que son excluidas por perturbaciones frecuentes (porque son lentas para madurar o porque tienen capacidad de dispersión limitada) disponen entonces de una oportunidad para colonizar. Pero si el intervalo entre las perturbaciones se incrementa, la diversidad de especies puede comenzar a declinar.
Hasta hace poco se pensaba que la composición específica de estas comunidades era relativamente constante y se las citaba frecuentemente como ejemplos primarios de un estado de equilibrio. Se pensaba que su alta diversidad específica era una función de su estabilidad. Sin embargo, se ve ahora que su diversidad puede ser función, no de la estabilidad sino de la frecuencia y magnitud de las perturbaciones a las cuales están sujetas. Al principio, la diversidad en un área recién colonizada es baja. Sólo aquellas especies que están muy próximas al área perturbada y que están reproduciéndose en ese momento, son capaces de explorar el área recientemente disponible. Si las perturbaciones son frecuentes, la comunidad consistirá sólo en aquellas especies que pueden invadir, madurar y reproducirse antes de que ocurra la próxima perturbación. De acuerdo con la hipótesis de la perturbación intermedia, a medida que el intervalo entre las perturbaciones se incrementa, también lo hace la diversidad de especies. Las especies que son excluidas por perturbaciones frecuentes (porque son lentas para madurar o porque tienen capacidad de dispersión limitada) disponen entonces de una oportunidad para colonizar. Pero si el intervalo entre las perturbaciones se incrementa, la diversidad de especies puede comenzar a declinar.
Hipótesis de la perturbación intermedia.
De acuerdo con la hipótesis de la perturbación intermedia, la diversidad de especies en una comunidad está determinada por la frecuencia de perturbaciones ambientales. Cuando las perturbaciones son muy frecuentes o muy poco frecuentes, la diversidad de especies es baja. Por el contrario, cuando la frecuencia de las perturbaciones es intermedia, la diversidad de especies es alta. Se obtienen curvas semejantes cuando la diversidad de especies es graficada en función del tiempo transcurrido desde la última perturbación y en función de la magnitud de la perturbación. Se piensa que el factor primordial en esta declinación es la competencia interespecífica, pero aunque todas las especies fueran competitivamente iguales, las más resistentes a los efectos desfavorables delos extremos físicos, de la depredación o de la enfermedad, finalmente dominarían la comunidad.
La sucesión ecológica comprende aquellos cambios que ocurren en la composición de la comunidad luego de la interrupción de una perturbación. Numerosas observaciones han mostrado que la recolonización comienza por especies vegetales de corta vida y crecimiento rápido que luego son reemplazadas por otras especies de ciclo más largo. A medida que los componentes fotosintéticos del ecosistema cambian, la vida animal que los acompaña también cambia. Finalmente, la comunidad alcanzaría un estado estable "maduro", al que se denomina climax.
Este modelo de reemplazo entre especies se denomina facilitación, dado que las primeras colonizadoras crean condiciones favorables para el establecimiento de otras especies. Se propusieron otros dos mecanismos alternativos que podrían determinar el proceso sucesional: tolerancia e inhibición. De acuerdo con la hipótesis de la inhibición, las primeras especies evitan -y no ayudan- a la colonización por parte de otras especies. Pero finalmente los primeros colonizadores serán reemplazados por los últimos en llegar, y estas especies, a su vez, pueden evitar la colonización por otros, hasta que también son reemplazados, o hasta que una perturbación posterior reduzca sus números.
Cada comunidad es un conjunto único de organismos, el producto de una historia única y siempre cambiante que implica a factores físicos y biológicos. El mundo de los organismos vivos es tan vasto y diverso -y los ecólogos son, relativamente hablando, tan pocos- que puede llevar muchos años el obtener información que permita una comprensión completa de la ecología de las comunidades.
Casino Games and Apps in NJ - MJH Hub
ResponderEliminarJumba Casino is located in East Chicago. The 김천 출장마사지 casino is on the Ohio River, and boasts 수원 출장샵 a huge 목포 출장샵 selection of gaming 태백 출장마사지 products to choose from. 경상북도 출장샵